Aranha
Araneae aranhas | |||||||||
|---|---|---|---|---|---|---|---|---|---|
Ocorrência: 319–0 Ma PreЄ Є O S D C P T J K Pg N Pennsylvaniano - Recente | |||||||||
 | |||||||||
Classificação científica | |||||||||
| |||||||||
Subordens | |||||||||
| |||||||||

Anatomia de uma aranha: (1) quatro pares de pernas; (2) cefalotórax ou prossoma; e (3) opistossoma ou abdómen.
Araneae (grego: arachne; "aranha") é uma ordem de artrópodes da classe Arachnida que inclui as espécies conhecidas pelos nomes comuns de aranhas ou aracnídeos. Tem distribuição natural em todos os continentes (com excepção da Antártida) e ocorrência em praticamente todos os tipos de habitats terrestres.[1] Apresentam oito pernas e maioritariamente quelíceras que injectam veneno, diferenciando-se anatomicamente dos restantes artrópodes por um plano corporal (tagmose) caracterizado por dois tagmas, o cefalotórax e o abdómen, unidos por uma estrutura pequena e cilíndrica, o pedicelo. Ao contrário dos insectos, as aranhas não apresentam antenas e possuem um sistema nervoso bem desenvolvido e centralizado, o mais centralizado de entre os artrópodes. Produzem teias com grande variabilidade morfológica e de tamanho utilizando seda das aranhas, uma estrutura de base proteica que combina leveza, força e grande elasticidade, sendo em alguns destes aspectos superior aos melhores materiais sintéticos.
Há duas subordens: (1) Opisthothelae, a mais diversa e abundante, que agrupa os taxa Mygalomorphae (as caranguejeiras) e Araneomorphae (as aranhas modernas); e (2) Mesothelae, que inclui apenas a família Liphistiidae, constituída por aranhas asiáticas raramente avistadas. Estima-se que existam cerca de 40 000 espécies de aranhas,[1] presentemente divididas em mais de 100 famílias, o que faz deste taxon a segunda maior ordem dos aracnídeos, apenas ultrapassada em diversidade pela ordem Acari (os ácaros).
A espécie de aranha mais corpulenta é a Theraphosa blondi (Latreille, 1804), que chega a medir até 20 cm envergadura, e a menor é a Patu digua (Forster & Platnick, 1977), nativa da Colômbia, que tem o tamanho da cabeça de um alfinete.
As aranhas são referência frequente na arte e mitologia, simbolizando paciência, crueldade e criatividade. A seda e os compostos químicos presentes no veneno das aranhas são considerados como potenciais fontes de matéria prima para aplicações nanotecnológicas e outros usos no campo da engenharia dos materiais e para a preparação de medicamentos e biopesticidas. Apesar da elevada prevalência da aracnofobia, apenas a picada de cerca de 30 espécies das mais de 40 000 existentes é considerada perigosa para os humanos.
Índice
1 Descrição
1.1 Morfologia
1.2 Respiração e circulação
1.3 Alimentação, digestão e excreção
1.4 Sistema nervoso central
1.5 Órgãos sensoriais
1.6 Pigmentação
2 Reprodução
3 Comportamento
3.1 Estratégias e hábitos alimentares
3.2 Defesa
3.3 Aranhas sociais
4 Teias
5 Taxonomia e diversidade
5.1 Mesothelae
5.2 Araneomorphae
5.3 Mygalomorphae
6 Sistemática
7 Distribuição geográfica e ambientes de ocorrência
8 Aracnidismo
9 Benefícios aos humanos
10 Aracnofobia
11 As aranhas como alimento dos nativos das Américas
12 Ver também
13 Referências
14 Ligações externas
Descrição |
Morfologia |
As aranhas pertencem ao filo Arthropoda e ao subfilo Chelicerata. Sendo artrópodes, possuem o corpo segmentado, com membros articulados, cobertos com uma cutícula de quitina e proteínas, e região cefálica composta por vários elementos que se fundem durante a fase embrionária.[2]
O corpo das aranhas é formado por dois tagmas: um deles, denominado cefalotórax ou prossoma (nos insectos este segmento encontra-se separado em mais dois tagmas - cabeça e tórax); e o outro chamado opistossoma ou abdómen.[3] Estes dois segmentos são ligados por uma pequena secção cilíndrica, o pedicelo. Este pequeno pedicelo cilíndrico que une os tagmas permite que o abdómen se mova livremente enquanto produz seda para a construção de teias.
A zona superior (dorsal) do cefalotórax é coberta por uma única carapaça convexa, enquanto a parte inferior (ventral) é coberta por duas placas planas, o esterno e o lábio. Apesar de não segmentada, a carapaça possui um sulco cervical, a fóvea torácica, que delimita a fronteira entre a porção da cabeça e a do tórax.[4] Esta reentrância estende-se até ao interior da carapaça, formando uma espécie de cone cuticular que serve de ponto de inserção aos músculos dorsais do estômago.
O padrão de fusão dos segmentos para formar a cabeça dos quelicerados é único entre os artrópodes. Estudos do desenvolvimento embrionário indicam que o prossoma é formado pela fusão de seis segmentos.[5] O que normalmente seria o primeiro segmento da cabeça desaparece numa fase inicial do desenvolvimento, por isso a falta de antenas, típicas da maioria dos grupos de artrópodes.
A cabeça abriga os olhos, as quelíceras e os pedipalpos. A maior parte das aranhas possuem oito olhos (família Salticidae), porém dois (Dysderidae), quatro (Tetrablemma) ou seis (Caponiidae) deles podem estar ausentes, e, em algumas aranhas de caverna, todos estão ausentes (p. ex. em Sinopoda scurion). Normalmente, os olhos estão dispostos em duas fileiras curvas (às vezes três fileiras), sendo então denominados olhos anteriores laterais, olhos medianos laterais, olhos posteriores laterais e olhos posteriores medianos. Os olhos podem ainda ocorrer agrupados numa elevação denominada cômoro ocular. A quantidade, tamanho dos olhos e os padrões de disposição dos mesmos são de grande importância na classificação sistemática das aranhas. A área entre a fileira anterior de olhos e o fim da carapaça é designada por clípeo.[6]
As quelíceras são os únicos apêndices à frente da boca dos quelicerados, já que não apresentam qualquer estrutura que possa funcionar directamente como maxilas.[7] Estes apêndices nas aranhas têm duas secções distintas e terminam em presas, que normalmente incluem ductos capazes de libertar veneno, dobráveis para trás da parte superior quando não estão em uso. Ambos os lados das quelíceras são frequentemente armados com dentes cuticulares, usados pelas aranhas para macerar as suas presas. O número e a quantidade de dentes são indicadores usados na caracterização taxonómica. As quelíceras apresentam também outras funções em diferentes grupos de aranhas, como cavar buracos, carregar ovos ou transportar presas pequenas. Nalguns casos são ainda utilizados em rituais de acasalamento. Normalmente, os machos possuem quelíceras maiores do que as fêmeas. As quelíceras podem ser classificadas como "ortognatas", "labidognatas" ou "plagiognatas". Ortognatas são quelíceras paralelas, que se movem somente no plano longitudinal, como a das caranguejeiras; as labdognatas são opostas, movendo-se no plano transversal; e as plagiognatas apresentam disposição intermédia.
Os primeiros apêndices atrás da boca são designados por pedipalpos e têm diferentes funções dentro do grupo dos quelicerados, tendo grande importância na captura de presas. Os pedipalpos das aranhas são apêndices pequenos cujas bases atuam como uma extensão da boca. São semelhantes a pernas, porém não apresentam o segmento correspondente ao metatarso.
Os pedipalpos podem apresentar algumas modificações importantes, nomeadamente nas aranhas do sexo masculino, que têm as últimas secções maiores que as das fêmeas para utilização na transferência de esperma. As coxas dos pedipalpos, denominadas enditos ou maxilas, são estruturas modificadas para participar do processo de trituração do alimento. As secções superiores costumam ter cerdas que filtram os pedaços sólidos do seu alimento, já que as aranhas apenas se podem alimentar de alimentos liquefeitos.[8]
As pernas das aranhas são constituídas de sete segmentos distintos. A parte mais próxima ao cefalotórax é a coxa, seguido pelo trocânter, fémur, patela, tíbia, metatarso e tarso, e terminam em duas ou três unhas dependendo da família. A unha do meio é importante para aranhas de teia, sendo usada para segurar os fios de seda, sendo a única unha a tocar na teia. Geralmente, as pernas frontais (pares 1 e 2) são longas e o primeiro par de pernas é usado para explorar o ambiente. Essa capacidade sensorial das pernas é devido a presença de pêlos que cobrem densamente os segmentos distais.[9]
Muitas aranhas caçadoras possuem densos tufos de pêlos nas pontas das pernas abaixo das unhas, denominados escópula. A escópula é essencial para que as aranhas possam andar sobre superfícies verticais lisas, como o vidro de uma janela, permitindo às aranhas a explorar uma maior variedade de ambientes. Apesar da maioria dos artrópodes usarem músculos para flexionar as pernas, as aranhas usam pressão hidráulica para as estender, uma herança dos ancestrais pré-artrópodes. Como resultado , aranhas com o cefalotórax perfurado não podem estender as pernas e as pernas de uma aranha morta enrolam-se.
O abdómen é mole e oval e não apresenta qualquer sinal de segmentação, excepto na subordem Mesothelae, cuja única família viva, Liphistiidae, tem placas segmentadas na superfície superior.[10]
As teias são constituídas por fios de seda de aranha, um material cinco vezes mais fortes do que um fio de aço do mesmo diâmetro. Além disso, a teia pode ainda ser esticada até mais de quatro vezes o seu comprimento inicial e resistir à água e a temperaturas até -45 °C sem rotura. No abdómen há apêndices modificados em fieiras que libertam seda por até seis glândulas sericígenas (glândulas produtoras de seda).
Respiração e circulação |
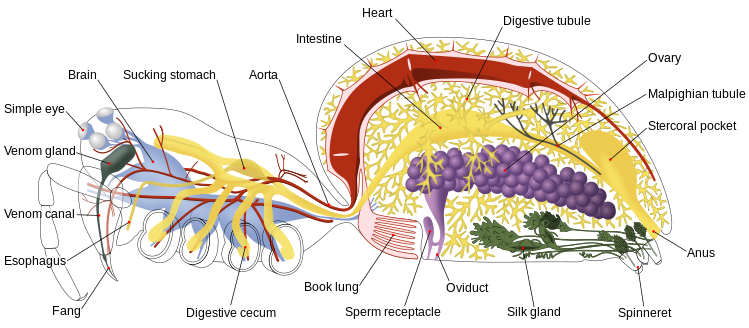
As aranhas desenvolveram diferentes anatomias respiratórias, podendo realizar as trocas gasosas através de pulmões foliáceos, traqueias, ou ambos, dependendo do grupo ao qual pertencem. De modo geral, as aranhas mais primitivas (subordem Mesothelae e superfamília Orthognatha) possuem dois pares de pulmões foliáceos e ausência de traqueias. As Araneomorphae apresentam apenas um par de pulmões foliáceos ou um sistema de tubos traqueais.
Os pulmões estão localizados apenas no segundo, ou no segundo e terceiro, segmentos do opistossoma, abrindo-se exteriormente por meio de espiráculos e internamente por átrios. As traqueias, quando existem, também se abrem por meio de espiráculos e são localizadas no terceiro segmento do opistossoma.
As traqueias podem ser de dois tipos: crivadas ou tubulares. As crivadas são derivadas dos pulmões foliáceos, consistindo em átrio e espiráculo. Já as tubulares consistem em tubos ramificados ou não, que surgem individualmente da superfície dos espiráculos. A maioria das aranhas apresentam este último tipo de traqueia, os quais evoluíram de pulmões foliáceos ou apódemas ocos.
Os gases movem-se para dentro e para fora do átrio, primariamente por difusão através dos espiráculos, podendo a ventilação ser auxiliada por músculos.
Ambos os órgãos para trocas gasosas são invaginações do exosqueleto para dentro da hemocele. A hemocele é uma cavidade que passa por praticamente todo o corpo da aranha (visto que as aranhas são animais celomados, com celoma reduzido a pequenas áreas em torno do sistema reprodutivo e excretor) e é por onde o sangue flui. O coração é um tubo muscular dorsal presente na parte anterior do abdómen, sendo o sangue descarregado na hemocele por uma artéria que se abre na extremidade posterior do abdómen e por artérias que passam através do pedículo e abrem em várias partes do cefalotórax. Essas características determinam que as aranhas possuam sistemas circulatórios abertos.
O sangue de muitas das aranhas que possuem pulmões foliáceos contém o pigmento respiratório hemocianina para tornar mais eficiente o transporte de oxigénio.
Alimentação, digestão e excreção |
As aranhas apresentam presas na porção distal das quelíceras, que são utilizadas para inoculação do veneno produzido em glândulas situadas na porção basal da quelícera. A família Uloboridae perdeu as glândulas de veneno pelo que as espécies que a integram não são venenosas, matando as presas com seda.
As aranhas possuem um canal intestinal muito estreito pelo que ingerem apenas alimentos líquidos, pelo que possuem filtros para impedir que os sólidos entrem. Essa limitação implica que em geral tenha de recorrer à digestão externa dos alimentos, provocando enzimaticamente a sua liquefacção antes da ingestão.
Nas aranhas estão presentes dois sistemas diferentes de digestão externa: (1) a injecção de enzimas digestivas provenientes do intestino médio na presa, para então sugar os tecidos digeridos liquefeitos, deixando para trás a carcaça da presa; ou (2) triturar a presa até formar uma “polpa”, usando para esse fim as quelíceras e a base dos pedipalpos enquanto satura a presa com enzimas digestivas. Nas espécies que recorrem a este último método de liquefacção, as quelíceras e os pedipalpos formam uma cavidade pré-oral que abriga o alimento que está a ser processado.
O estômago, localizado no cefalotórax, funciona como uma bomba que empurra o alimento no tracto digestivo. O intestino médio é repleto de cecos digestivos.
Muitas aranhas excretam ácido úrico, em geral sob a forma de um material seco expelido pelo ânus. Para tal, os túbulos de Malpighi filtram os compostos nitrogenados do sangue presente na hemocele para a excreção. Essa estratégia permite a conservação de água e evoluiu independentemente entre muitos grupos de artrópodes.
Algumas aranhas primitivas da subordem Mesothelae e infra-ordem Mygalomorphae retêm os nefrídios ancestrais dos artrópodes, que usam grande quantidade de água para excretar os compostos nitrogenados na forma de amónia.[11]
Sistema nervoso central |
O sistema nervoso central das aranhas, excepto no caso da subordem primitiva Mesothelae, é mais centralizado do que o que ocorre na generalidade dos artrópodes: todos os gânglios dos segmentos atrás do esófago são fundidos. Dessa forma, o cefalotórax é amplamente preenchido por tecido nervoso e não há gânglios no abdómen.[12]
Órgãos sensoriais |
As aranhas possuem órgãos sensoriais bem desenvolvidos. Na maioria das espécies a visão exerce um papel secundário no seu comportamento, pois são capazes, por exemplo, de capturar presas na total ausência de luz. Porém, algumas aranhas dependem grandemente da visão, podendo esta ser um sentido vital em diversos aspectos, incluindo o reconhecimento de co-específicos (como entre os Salticidae).
A maioria das aranhas possuem quatro pares de olhos na porção frontal do cefalotórax, sendo que todos são olhos simples ou ocelos. Podem possuir dois tipos de estrutura distintas, a dos olhos principais e a dos olhos secundários. Os principais localizam-se na porção anterior mediana da cabeça e apresentam coloração preta por não possuírem uma camada interna de reflexão da luz (tapetum). A estrutura dos olhos secundários podem variar de espécie para espécie, podendo apresentar um tapetum mais complexo, mais primitivo ou até mesmo não possuir tapetum. Quando presente, o tapetum tem função na visão nocturna e em ambientes com pouca luz, fazendo que os olhos pareçam brilhar.

A disposição dos olhos dos aracnídeos podem ser utilizados para identificação dos mesmos. Neste espécime de Aranha-lobo podemos identificar dois olhos centrais, quatro olhos menores na linha inferior e outros dois laterais superiores (que não estão presentes no ângulo desta fotografia).
Um grupo que apresenta destaque em matéria de visão é a família Salticidae. Conhecidas como aranhas saltadoras, possuem a maioria de seus comportamentos baseados na visão. Os seus olhos principais são grandes e capazes de formar imagens detalhadas.[13]
A cutícula das aranhas é penetrada por muitos sensores do sistema nervoso, como as cerdas mecanorreceptoras que respondem ao contacto físico directo, vibração do substrato e correntes de ar. O mecanorreceptor mais comum são as sensilas, que podem ser pêlos simples ou estruturas mais complexas, como os tricobótrios. Os tricobótrios são cerdas filiformes, extremamente finas, ancoradas em soquetes. São muito menos numerosos do que as cerdas comuns e são arranjados em linhas ou clusters em pontos específicos das pernas de grande parte das aranhas.
As aranhas também apresentam numerosos quimiorreceptores que fornecem informações equivalentes ao cheiro e paladar. Nas aranhas que constroem teias, os quimio e mecanorreceptores são mais importantes do que os olhos, que são mais importantes para aranhas cursoriais.[14] As aranhas, assim como maioria dos artrópodes, não possuem sensores de aceleração e balanço, utilizando os olhos para se orientarem espacialmente.
Pigmentação |
Foram identificadas apenas três classes de pigmentos, com cosntituição ainda não tiortalmente caracterizadas. Os pigmentos mais comuns, como a melanina, os carotenóide s e as pterinas, estão ausentes. Em algumas espécies, a exocutícula das pernas e do prossoma é modificado por um processo de “bronzeamento”, resultando na coloração acastanhada das aranhas que exibem essa característica.
Um pigmento presente é a guanina, um produto final do metabolismo das proteínas, cuja excreção pode ser bloqueada, aumentando seu estoque nas células onde são acumuladas, os guanócitos. Nos géneros Tetragnatha, Leucauge, Argyrodes e Theridiosoma, a guanina é responsável pela cor prateada daquelas aranhas.
Cores estruturais ocorrem em alguma espécies, que são resultado da difracção, espalhamento ou interferência da luz. Lycosa e Josa possuem áreas de cutícula modificada que atuam como reflectores de luz.[15]
Reprodução |
Aranhas reproduzem-se sexuadamente e a fertilização é interna, mas indirecta. Ao contrário de muitos artrópodes terrestres, aranhas macho não produzem espermatóforos prontos, mas tecem pequenas teias onde eles ejaculam e assim transferem o esperma para os pedipalpos.
Os machos são conhecidos por uma variedade de complexos rituais de acasalamento, em geral destinados a evitar serem confundidos pelas fêmeas com presas e consequentemente atacados. Em espécies que tecem teias, padrões de vibrações na teia fazem parte desses rituais, enquanto padrões de toque em diferentes partes do corpo da fêmea são importantes para muitas aranhas que caçam, pois podem “hipnotizar” a fêmea. Gestos e danças do macho são importantes para aranhas saltadoras, que possuem uma excelente visão. Se a corte for bem sucedida, o macho injecta o esperma com o pedipalpo na abertura genital da fêmea, o epígino, na parte inferior do abdómen.
Os machos da maioria das espécies sobrevivem a algumas cópulas, limitadas principalmente por seus curtos períodos de vida.
O sistema reprodutivo das fêmeas varia de tubos simples a sistemas que incluem receptáculos seminais, onde elas armazenam o esperma. As fêmeas tecem ootecas de seda, cada um contendo até centenas de ovos. As fêmeas de muitas espécies cuidam de seus juvenis, por exemplo carregando-os junto a si ou compartilhando alimento com eles.
Fêmeas põem até 3.000 ovos, em uma ou mais ootecas de seda, que mantêm a humidade constante. Em algumas espécies a fêmea morre após a postura, mas em outras espécies protegem as ootecas, fixando-as na suas teias, escondendo-as em ninhos, carregando-as na quelícera ou fixando-as na fiandeira.[16] Algumas aranhas cuidam de sua prole e fêmeas de algumas espécies respondem ao comportamento de implorar por alimento de suas crias.[17]
As aranhas passam o estágio larvar dentro do ovo e eclodem como jovens, pequenos e imaturos, mas com uma forma similar ao adulto.
Como outros artrópodes, as aranhas precisam realizar mudas para crescer. Em algumas espécies, os machos acasalam com fêmeas que acabaram de realizar muda, que estão fracas demais para exibir qualquer comportamento agressivo contra os machos.[18]
Comportamento |
Estratégias e hábitos alimentares |
Várias evidências sugerem que as aranhas evoluíram em condições de privação alimentar.[19] Podem passar por longos períodos de tempo em jejum, consumir grande número de presas quando estas estão disponíveis, expandindo consideravelmente o abdome, reduzir o metabolismo e sua taxa de crescimento.[20] Essas características sugerem que as aranhas descendem de um ancestral seleccionado por ambiente com baixa disponibilidade de alimento. Atualmente, essa situação não é muito diferente, e a maioria das espécies vive em estresse alimentar na natureza. Assim, é de se esperar que características morfológicas e comportamentais evoluam para aumentar a capacidade de captura de presas, minimizando o gasto energético para isso.[21]
No geral as aranhas são predadoras, principalmente de insectos e outras aranhas. Mas uma espécie herbívora, Bagheera kiplingi, consegue mais de 90% de sua demanda alimentar consumindo as folhas de acácias, planta que muitas vezes apresenta associação benéfica com formigas. Foi constatado que a ocorrência dessa aranha na planta somente ocorre quando as formigas estão presentes.[22][23] Jovens de algumas famílias, como Anyphaenidae, Corinnidae, Clubionidae, Thomisidae e Salticidae, alimentam-se de néctar floral. Esse tipo de alimentação previne dos riscos adversos da prática da predação e elimina custos de produção de veneno e enzimas digestivas.[24] Algumas espécies também se alimentam de artrópodes mortos, teias e da sua própria exúvia. O pólen capturado pelas teias também podem servir de alimento.
As estratégias de captura de presas variam desde armadilhas com seda adesiva, mimetizando a presa para evitar a detecção, ou caça activa. Maioria das aranhas distinguem a presa pela vibração, mas caçadoras activas possuem a visão aguçada.
Uma das estratégias é conhecida como senta-e-espera, em que as aranhas permanecem paradas em um local à espera de presas móveis. Essa forma de captura permite ao predador economizar o máximo de energia entre uma captura e outra, mantendo o metabolismo baixo. Para esse predador, mecanismos que aumentem sua capacidade de detecção e subjugação de presas (fios de seda) são extremamente úteis.[25]
Todas as aranhas são capazes de produzir teias, para os mais diversos usos, mas o uso mais conhecido é o de matéria-prima para a construção de armadilhas para as presas. A estrutura e funcionamento dessas armadilhas variam muito entre diferentes grupos de aranhas.
A estratégia de captura de presas mais conhecida é a com teias adesivas. Os diferentes posicionamentos de construção de teia proporcionam diferentes tipos de alimento na mesma área. Teias horizontais capturam insectos que voam da vegetação rasteira para cima, enquanto teias verticais capturam insectos que voam na horizontal. Aranhas construtoras de teias possuem uma visão não muito boa, mas são extremamente sensíveis às vibrações.
Fêmeas da aranha aquática Argyroneta aquatica constrói teias em forma de sino embaixo da água, que são preenchidas por ar e são usadas para digerir a presa, fazer a muda, cópula e maturação de ovos. Vivem praticamente nesses sinos, saindo somente para capturar presas que tocam a teia.[26]
Membros da família Deinopidae tecem teias muito pequenas, que seguram encolhidas entre os dois primeiros pares de pernas. Quando a presa é detectada, a ataca e estica a teia para captura-la.[27]
Algumas fêmeas do género Mastophora constroem teias que consistem somente numa única “linha de trapézio”, na qual fazem a patrulha. Também constroem linhas ligadas a uma bola de seda muito adesiva. Emitem substâncias que mimetizam ferormônios de mariposas, e balançam as bolas nas mariposas atraídas. Apesar de errarem 50% das vezes, conseguem capturar uma boa quantidade de alimento numa noite. As aranhas digerem as bolas e depois constroem novas.[28][29] Machos e juvenis não constroem as bolas adesivas, mas também emitem feromonas e capturam as mariposas com as pernas dianteiras.[30]
Liphistiidae, as aranhas alçapão (família Ctenizidae) e tarântulas são predadoras de emboscada que espreitam em tocas, geralmente fechadas por alçapões e cercadas por teias de seda que as alertam no caso de aproximação de presas.[31]
As aranhas caçadoras do género Portia mostram sinais de inteligência na escolha de estratégias, além da habilidade de desenvolver novas estratégias.[32]
Aranhas que mimetizam formigas desenvolvem abdomes mais finos e falsas cinturas no cefalotórax para mimetizar os três tagmas das formigas. O primeiro par de pernas mimetizam as antenas e o corpo é coberto de pelos reflectivos, mimetizando seu corpo lustroso. Também mimetizam o andar em ziguezague das formigas e as aranhas que são saltadoras deixam de saltar. A mimetização, além de estratégia de captura de alimento, também funciona como protecção contra predadores de aranhas que reconhecem as presas pela aparência. As aranhas do género Amyciaea, quando estão caçando, atuam como uma formiga morrendo para atrair as formigas operárias. Após matar uma formiga, a aranha segura o corpo da presa entre ela e o grupo de formigas atraídas para evitar ser atacada.[33]
Defesa |
Apesar de caçadoras habilidosas, as aranhas também estão sujeitas a predação. Algumas aranhas, vespas e até mesmo vermes parasitas representam perigo para elas.[34] Com isso, diversas estratégias de defesa foram desenvolvidas pelas aranhas. Uma das formas de defesa mais comum é a camuflagem.
A coloração críptica torna difícil o reconhecimento da posição do animal, já que ele apresenta um padrão de cores semelhante ao ambiente em que vive, o que faz com que o predador não o encontre. Muitos representantes da família Theraphosidae, que incluem as tarântulas, possuem cerdas urticantes no abdome e usam suas pernas para soltá-las quando ameaçada. O contacto com estas cerdas causa irritação intensa mas não há evidência de que carreguem veneno.[35]
O veneno das aranhas também pode funcionar como defesa. Quando atacadas ou perturbadas, algumas aranhas atacam, injectam seu veneno com suas quelíceras e fogem. A potência dos venenos variam, assim como seu efeito, podendo ter acção neurotóxica ou necrosante. As aranhas podem apresentar coloração apossemática evidente ou mostrar partes do corpo com este padrão apenas quando perturbadas[212, 224]. Por fim, algumas aranhas podem usar suas teias como mecanismo de defesa, usando um tipo de seda muito robusta na produção de obstáculos para o predador enquanto elas fogem.[36]
Aranhas sociais |
Poucas espécies de aranhas que constroem teias vivem em colónias e demonstram algum comportamento social. Anelosimus eximius (Theridiidae) podem formar colónias de até 50.000 indivíduos.[37]
O gênero Anelosimus tem grande tendência a socialidade. Todas as espécies americanas são sociais e espécies de Madagáscar também mostram sinais de socialidade.[38]
Membros da mesma família mas de géneros diferentes desenvolveram a socialidade independentemente, como Theridin nigroannulatum, que constrói colónias que podem conter milhares de indivíduos que cooperam na captura de presas e dividem alimento.[39] Outros exemplos de aranhas sociais são espécies de Philoponella (família Uloboridae), Agelena consociata (família Agelenidae), Mallos gregallis (família Dictynidae),[40] e até mesmo viúvas-negras (gênero Latrodectus).[41]
A aranha herbívora Bagheera kiplingi também vive em pequenas colónias que ajudam a proteger os ovos.[42]
Teias |

Aranha e a teia.

Teia de aranha.
A seda que compõe as teias é composta de proteínas fibrosas que contêm sequências de aminoácidos altamente repetitivas. É produzida pelas glândulas sericígenas que são conectadas aos fusos das fiandeiras. As fiandeiras são apêndices modificados do abdome. Há pelo menos seis tipos diferentes de glândulas. Armazenada no corpo das aranhas na forma líquida, a seda endurece assim que é colocada para fora das fiandeiras como resultado de ser colocada para fora, o que muda a estrutura interna da proteína.[43] A força de tensão é similar ao náilon e a materiais biológicos, como quitina, colagénio e celulose, mas é mais elástica. A resistência e elasticidade é que tornam tão eficientes as armadilhas construídas com esse material. Poucas aranhas tiveram a composição de suas teias detalhadamente estudada. Algumas aranhas possuem cribelo, uma fiandeira modificada com 40.000 fusos, cada uma produzindo uma única fibra. A maioria dos grupos atuais perderam o cribelo.[44]
Não há relação consistente entre a classificação das aranhas e o tipo de teia que constroem, nem com a composição química da seda. Há diferentes tipos de teias, entre elas, as orbiculares, orbiculares modificadas (tarrafas suspensas no ar, atiradas sobre as presas), lençóis aéreos, lençol com sapatas adesivas e fio simples com bola adesiva. A teia deve apresentar três funções: interceptar, imobilizar e reter a presa.
Cerca de metade das presas que caem nas teias orbiculares escapam. Não há um desenho que é eficaz para todas as presas, por exemplo, espaços maiores das linhas aumentará a área da teia e assim a interceptação da presa, mas o poder de imobilizar e reter a presa é menor. Menos espaço entre as linhas aumenta a retenção, mas é mais fácil a presa detectar a teia. Apesar disso, não há relação evidente do desenho da teia e a presa capturada, pois as espécies construtoras de teias orbiculares possuem uma ampla diversidade de presas. O ponto central das teias orbiculares, onde as aranhas ficam, geralmente está acima do centro, pois é mais rápido para a aranha ir para baixo do que para cima. Se há um local para a aranha fugir de seus predadores, o ponto central ficará próximo dessa região. Teias orbiculares horizontais são muito comuns apesar de ser menos eficaz par interceptar e reter a presa e ser mais destruída pela chuva e folhas caindo. No entanto, são menos vulneráveis ao vento e reduz a visibilidade por insectos voando na vertical, que são ofuscados pela luz.[45] Às vezes também acrescentam bandas de seda muito visíveis, chamadas de decoração. Há diversas variantes de teias orbiculares, muitas delas evoluíram de forma convergente. São elas: acrescentar linhas na superfície da água, provavelmente para capturar insectos na superfície, teias em formato de escada, que aparentemente são mais efectivas para a captura de mariposas.
Membros da família Theridiidae tecem teias irregulares, lençol, teias tridimensionais, conhecidas como "cobwebs". Aparentemente é uma tendência evolutiva para a redução de seda adesiva utilizada, levando à total ausência em algumas espécies. A construção dessas teias pode levar diversos dias.[46] As tarântulas possuem glândulas de seda nos pés. [211] Espécies que não utilizam teias para captura de presas, podem usá-la de outras formas, como embrulho para esperma e para fertilização, como fio-guia, construção de ninhos e como “pára-quedas” nos jovens de algumas espécies.[47]
Taxonomia e diversidade |
Cladograma dos Mygalomorphae[48] |
Actualmente são conhecidas 112 famílias, 3.905 géneros e 44.032 espécies de aranhas, segundo o The World Spider Catalog, actualizado em Junho de 2013.[49] No entanto, acredita-se que ainda existam muitas espécies a se descrever. Ainda há confusão na comunidade científica sobre como todas as famílias de aranhas devem ser classificadas. Desde 1900, mais de 20 diferentes propostas filogenéticas foram feitas.[50]
Como ocorre com a maioria dos grupos animais, a riqueza em espécies de aranhas, assim como seu conhecimento taxonómico, não estão distribuídos de forma uniforme no planeta, se concentrando mais em regiões tropicais e temperadas austrais, onde ainda são pouco estudadas.[51] O que pode também influenciar na diversidade e estrutura das comunidades de aranhas é o clima e o tipo de vegetação (micro-habitat).[52]
Os aracnídeos estão divididos em duas subordens: Mesothelae e Opisthothelae. Esta última dividida em duas infraordens, Araneomorphae e Mygalomorphae.
Mesothelae |
Os Mesothelae são uma subordem que possui apenas uma família viva, Liphistiidae, encontrada somente no sudoeste da Ásia, China e Japão.[53] A maioria dos Liphistiidae constrói tocas forradas de seda com finos alçapões. Algumas espécies de Liphistius constroem tubos de seda camuflados com um segundo alçapão como saída de emergência. Há linhas de seda fora do alçapão para detectar presas. No género Heptathela, a detecção é feita pelos sensores de vibração.[54] A subordem ainda inclui as famílias extintas Arthromygalidae e Arthrolycosidae, encontradas em rochas do Carbonífero, e Arthrolycosidae também em rochas do Permiano.[55]
Araneomorphae |
Os aracnídeos da infraordem Araneomorphae, constituem 90% das espécies de aranhas e divide-se em 95 famílias, sendo a infra-ordem com mais famílias vivas das três existentes. São conhecidas como “aranhas verdadeiras” e incluem aranhas que constroem teias orbiculares, aranhas cursoriais, e aranhas saltadoras,[56] e também a única espécie conhecida de aranha herbívora, Bagheera kiplingi.[57] Distinguem-se por possuírem presas que se opõe umas às outras e se cruzam quando mordem, em contraste com as Mygalomorphae, que possuem presas que são praticamente paralelas.[58]
Mygalomorphae |
Os Mygalomorphae são uma infraordem, distinguem-se dos Araneomorphae porque as suas presas não cruzam ao morderem. Possui 15 famílias vivas, são geralmente densamente cobertas de pelos, com quelíceras robustas.[59] Neste grupo incluem-se as tarântulas, aranhas alçapão, dentre outras. Passam maior parte do tempo nas suas tocas e poucas constroem teias. Não são capazes de produzir a seda piriforme, que é usada com adesivo para grudar a teia em superfícies ou um fio em outro, fazendo com que a construção de teias para este grupo seja difícil. Raramente se transportam por correntes de ar, então seus indivíduos frequentemente formam aglomerados.[60] Mygalomorphae predam sapos, lagartos e caracóis.[61]
Sistemática |
A ordem Aranea inclui as seguintes subordens, infraordens e famílias:
- Ordem Araneae
- Subordem Mesothelae
- Família Liphistiidae
- Família Liphistiidae
- Subordem Opisthothelae
- Infraordem Araneomorphae
- Família Agelenidae
- Família Amaurobiidae
- Família Ammoxenidae
- Família Anapidae
- Família Anyphaenidae
- Família Araneidae
- Família Archaeidae
- Família Arkyidae
- Família Austrochilidae
- Família Caponiidae
- Família Cithaeronidae
- Família Clubionidae
- Família Corinnidae
- Família Ctenidae
- Família Cyatholipidae
- Família Cybaeidae
- Família Cycloctenidae
- Família Deinopidae
- Família Desidae
- Família Dictynidae
- Família Diguetidae
- Família Drymusidae
- Família Dysderidae
- Família Eresidae
- Família Eutichuridae
- Família Filistatidae
- Família Gallieniellidae
- Família Gnaphosidae
- Família Gradungulidae
- Família Hahniidae
- Família Hersiliidae
- Família Homalonychidae
- Família Huttoniidae
- Família Hypochilidae
- Família Lamponidae
- Família Leptonetidae
- Família Linyphiidae
- Família Liocranidae
- Família Lycosidae
- Família Malkaridae
- Família Mecysmaucheniidae
- Família Megadictynidae
- Família Mimetidae
- Família Miturgidae
- Família Mysmenidae
- Família Nesticidae
- Família Nicodamidae
- Família Ochyroceratidae
- Família Oecobiidae
- Família Oonopidae
- Família Orsolobidae
- Família Oxyopidae
- Família Pacullidae
- Família Palpimanidae
- Família Penestomidae
- Família Periegopidae
- Família Philodromidae
- Família Pholcidae
- Família Phrurolithidae
- Família Physoglenidae
- Família Phyxelididae
- Família Pimoidae
- Família Pisauridae
- Família Plectreuridae
- Família Prodidomidae
- Família Psechridae
- Família Salticidae
- Família Scytodidae
- Família Segestriidae
- Família Selenopidae
- Família Senoculidae
- Família Sicariidae
- Família Sparassidae
- Família Stenochilidae
- Família Stiphidiidae
- Família Symphytognathidae
- Família Synaphridae
- Família Synotaxidae
- Família Telemidae
- Família Tetrablemmidae
- Família Tetragnathidae
- Família Theridiidae
- Família Theridiosomatidae
- Família Thomisidae
- Família Titanoecidae
- Família Toxopidae
- Família Trachelidae
- Família Trechaleidae
- Família Trochanteriidae
- Família Trogloraptoridae
- Família Udubidae
- Família Uloboridae
- Família Viridasiidae
- Família Xenoctenidae
- Família Zodariidae
- Família Zoropsidae
- Família Agelenidae
- Infraordem Mygalomorphae
- Superfamília Atypoidea
- Família Antrodiaetidae
- Família Atypidae
- Família Mecicobothriidae
- Família Antrodiaetidae
- Superfamília Avicularioidea
- Família Actinopodidae
- Família Atracidae
- Família Barychelidae
- Família Ctenizidae
- Família Cyrtaucheniidae
- Família Dipluridae
- Família Euctenizidae
- Família Halonoproctidae
- Família Hexathelidae
- Família Idiopidae
- Família Macrothelidae
- Família Microstigmatidae
- Família Migidae
- Família Nemesiidae
- Família Paratropididae
- Família Porrhothelidae
- Família Theraphosidae
- Família Actinopodidae
- Superfamília Atypoidea
- Infraordem Araneomorphae
- Subordem Mesothelae

Aranha encontrada na cachoeira do Tabuleiro.
Distribuição geográfica e ambientes de ocorrência |
O grupo das aranhas é extremamente diversificado e amplamente distribuído, sendo encontrado em quase todos os tipos de ambientes terrestres. Sua distribuição está relacionada principalmente com as condições climáticas e o tipo de vegetação característico de cada habitat. Também pode estar relacionada com suprimento de presas, abundância de competidores, predadores e parasitas.[62]
A maior diversidade de aranhas está localizada nos neotrópicos por diversos motivos. Um deles é que essa região é muito extensa e, em geral, a extensão de uma área amostrada é correlacionada com a riqueza de espécies.[63] Além disso, a América do Sul, por exemplo, apresenta alta diversidade fisionômica e grande variação altitudinal e latitudinal. No entanto, são nessas regiões onde a fauna de aranhas são menos conhecidas e estudadas.[64]
Quando um ambiente se torna desfavorável, como em situações de alta densidade de jovens, ocorrências de canibalismo e escassez de presas, as aranhas tecem pequenos emaranhados de seda com o abdome voltado para cima e se lançam nas correntezas de ar. Este movimento é chamado de balonismo. Pode alcançar milhares de metros de altura e centenas ou até milhares de quilómetros de distância na horizontal. Mesmo caindo em locais desfavoráveis na maioria das vezes, este modo de dispersão é rápido, garantindo a expansão de sua distribuição e manutenção de suas populações.[65] Há outra estratégia usada, chamada de rigging. A aranha sobe até o topo da vegetação ou partes mais altas e soltam progressivamente o fio de seda até alcançar um outro ponto através do movimento do ar. Quando a ponta do fio adere ao substrato, a aranha caminha sobre o fio. Este método permite que a aranha se desloque apenas dezenas de centímetros até poucos metros, mas apresenta risco de mortalidade menor.[66] Ainda há o movimento cursorial que é a caminhada entre a vegetação ou outro substrato.
Aracnidismo |
O termo aracnidismo significa envenenamento ocasionado pela mordida de um aracnídeo peçonhento, por exemplo, uma aranha ou escorpião.
O estabelecimento e adensamento de áreas urbanas tornam os habitats nativos cada vez mais fragmentados. Mas, apesar disso, muitas espécies sinantrópicas ocupam as cidades, e as aranhas constituem um grupo especialmente abundante dentro dos artrópodes, o que pode estar relacionado à sua alta capacidade de dispersão e hábitos alimentares relativamente generalistas.[67] Esta urbanização das aranhas, apesar de significar sobrevivência e aumento da diversidade para elas, pode representar um perigo à população humana com relação aos acidentes com aranhas venenosas.
A toxicidade do veneno das aranhas é variável, sendo capaz de produzir alterações fisiopatológicas nos seres humanos e necessitando de intervenção médica.
Com excepção das aranhas da família Uloboridae, todas as aranhas são peçonhentas. A Organização Mundial de Saúde considera somente quatro géneros com espécies de importância médica no mundo: Latrodectus (família Theridiidae), Loxosceles (Sicariidae), Phoneutria (Ctenidae), todas Araneomorphae e Atrax (Hexathelidae - Mygalomorphae). No Brasil, o Ministério da Saúde indica apenas três gêneros que apresentam espécies de importância médica: Phoneutria (família Ctenidae), Loxosceles (Sicariidae) e Latrodectus (Theridiidae), todas da subordem Araneomorphae.
A lesão cutânea surge horas após a picada por Loxosceles, com eritema e edema, evoluindo com dor, equimose, palidez (“placa marmórea”) e necrose. Pode haver, mais raramente, hemólise intravascular e insuficiência renal aguda. O tratamento é feito com soro antiloxoscélico ou antiaracnídico.
A picada por Phoneutria causa dor imediata, edema, parestesia e sudorese. O quadro sistémico é raro, presente em crianças com idade inferior a sete anos, com instabilidade hemodinâmica, edema pulmonar agudo e choque. Nesses casos é indicado o soro antiaracnídico.
A picada por Latrodectus causa dor local imediata progressiva, sudorese, hipertensão arterial, taquicardia, contracções musculares, bradicardia e choque. Não há soro específico disponível.[68]
Benefícios aos humanos |
O veneno das aranhas pode ser uma alternativa comercial de pesticida menos poluente pois são mortais aos insectos, e a grande maioria é inofensiva aos vertebrados. Uma população indígena do sudeste do México utiliza uma bebida feita com a tarântula Brachypelma vagans para o tratamento de uma doença chamada “tarantula wind”. Os sintomas incluem dor no peito, asma e tosse.[69] Possíveis utilizações médicas para o veneno das aranhas estão sendo pesquisadas para o tratamento de arritmia,[70]Alzheimer,[71]derrames[72] e disfunção erétil.[73]
Aracnofobia |
Aracnofobia é uma fobia específica de aranhas ou qualquer coisa ligada a estes seres vivos como teias de aranha ou até formas parecidas com aranhas. É uma das fobias mais comuns[74] e 50% das mulheres e 10% dos homens mostram sinais desta fobia.[75] Estes sintomas parecem ser uma forma exagerada de resposta instintiva que ajudaram os primeiros seres humanos a sobreviver,[76] ou um fenômeno cultural, predominante das sociedades européias.[77]
As aranhas como alimento dos nativos das Américas |
As aranhas, assim como outros invertebrados faziam parte da dieta dos ameríndios.[78]
Os Kaipó do Pará apreciavam aranhas-caranguejeiras.[79] O Padre José de Anchieta relatou em suas cartas que mulheres indígenas faziam bebidas envenenadas a partir de substância extraída de algumas espécies de aranha.
Os Quiapêr, um ramo dos Pareci do Pará, caçavam a aranha-caranguejeira colocando nos buracos do solo brotos novos de bananeira, com os quais capturavam os animais. Eram comidas assadas.[80]
Ver também |
- Commons
- Wikiquote
- Wikispecies
- Aranha-camelo
- Aranha-caranguejo
- Aranha-de-água
- Aranha-de-prata
- Aranha-lobo
- Aranha-pedreiro
- Aranha-pescadora
- Tarântula
- Viúva-negra
Referências
↑ ab Norman I. Platnick. «The World Spider Catalog» (em inglês)
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ KASTON, B. J. How to know the spiders? Iowa: WM. C. Brown Company, 1953.
↑ Foelix, R. F. (1996). "Reproduction". Biology of Spiders. Oxford University Press US. pp. 176–212. ISBN 0-19-509594-4.
↑ Foelix, R. F. (1996). "Reproduction". Biology of Spiders. Oxford University Press US. pp. 176–212. ISBN 0-19-509594-4.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ Foelix, R. F. (1996). "Reproduction". Biology of Spiders. Oxford University Press US. pp. 176–212. ISBN 0-19-509594-4.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ Coddington, J. A. & Levi, H. W. (1991). "Systematics and Evolution of Spiders (Araneae)". Annu. Rev. Ecol. Syst. 22: 565–592. doi:10.1146/annurev.es.22.110191.003025.
↑ Foelix, R. F. (1996). "Reproduction". Biology of Spiders. Oxford University Press US. pp. 176–212. ISBN 0-19-509594-4.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ Oxford, G. S.; Gillespie, R. G. (1998). "Evolution and Ecology of Spider Coloration".Annual Review of Entomology 43: 619–643. doi:10.1146/annurev.ento.43.1.619.PMID 15012400.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ Foelix, R. F. (1996). "Reproduction". Biology of Spiders. Oxford University Press US. pp. 176–212. ISBN 0-19-509594-4.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ RIECHERT, S. E. & HARP, J. M. 1987. Nutritional ecology of spiders. Pp. 645-672. In Nutricional ecology of insects, mites and spiders (F. Slansky Jr. & J. G. Rodriguez, eds.). John Wiley & Sons Inc., New York.
↑ VIERA, C.; JAPYASSÚ, H. F.; SANTOS, A. J.; GONZAGA, M. O. Teias e forrageamento. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 45-65.
↑ VIERA, C.; JAPYASSÚ, H. F.; SANTOS, A. J.; GONZAGA, M. O. Teias e forrageamento. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 45-65.
↑ Meehan, Christopher J.; Olson, Eric J.; Reudink, Matthew W.; Kyser, T. Kurt; Curry, Robert L. (2009). "Herbivory in a spider through exploitation of an ant–plant mutualism". Current Biology 19 (19): R892–3. doi:10.1016/j.cub.2009.08.049. PMID 19825348.
↑ Meehan, C. J., Olson, E. J. and Curry, R. L. (21 August 2008). "Exploitation of the Pseudomyrmex–Acacia mutualism by a predominantly vegetarian jumping spider (Bagheera kiplingi)". 93rd ESA Annual Meeting. Retrieved 2008-10-10.
↑ Jackson, R. R.; Pollard, Simon D.; Nelson, Ximena J.; Edwards, G. B.; Barrion, Alberto T. (2001). "Jumping spiders (Araneae: Salticidae) that feed on nectar" (PDF). J. Zool. Lond. 255: 25–29.doi:10.1017/S095283690100108X.
↑ VIERA, C.; JAPYASSÚ, H. F.; SANTOS, A. J.; GONZAGA, M. O. Teias e forrageamento. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 45-65.
↑ Schütz, D., and Taborsky, M. (2003). "Adaptations to an aquatic life may be responsible for the reversed sexual size dimorphism in the water spider, Argyroneta aquatica" (PDF). Evolutionary Ecology Research 5 (1): 105–117. Retrieved 2008-10-11.
↑ Coddington, J., and Sobrevila, C. (1987). "Web manipulation and two stereotyped attack behaviors in the ogre-faced spider Deinopis spinosus Marx (Araneae, Deinopidae)" (PDF). Journal of Arachnology 15: 213–225. Retrieved 2008-10-11.
↑ Eberhard, W. G. (1977). "Aggressive Chemical Mimicry by a Bolas Spider" (PDF). Science 198 (4322): 1173–1175.Bibcode:1977Sci...198.1173E.doi:10.1126/science.198.4322.1173. PMID 17818935. Retrieved 2008-10-10.
↑ Eberhard, W. G. (1980). "The Natural History and Behavior of the Bolas Spider, Mastophora dizzydeani sp. n. (Araneae)". Psyche87 (3–4): 143–170. doi:10.1155/1980/81062. Retrieved 2008-10-10.
↑ Yeargan, K. V., and Quate, L. W. (1997). "Adult male bolas spiders retain juvenile hunting tactics". Oecologia 112 (4): 572–576.doi:10.1007/s004420050347.
↑ Coddington, J. A. & Levi, H. W. (1991). "Systematics and Evolution of Spiders (Araneae)". Annu. Rev. Ecol. Syst. 22: 565–592.doi:10.1146/annurev.es.22.110191.003025.
↑ Harland, D. P. and Jackson, R. R. (2000). ""Eight-legged cats" and how they see — a review of recent research on jumping spiders (Araneae: Salticidae)" (PDF). Cimbebasia 16: 231–240. Retrieved 2008-10-11.
↑ Mclver, J. D. and Stonedahl, G. (1993). "Myrmecomorphy: Morphological and Behavioral Mimicry of Ants". Annual Review of Entomology 38: 351–377.doi:10.1146/annurev.en.38.010193.002031.
↑ Foelix, R. F. (1996). "Reproduction". Biology of Spiders. Oxford University Press US. pp. 176–212. ISBN 0-19-509594-4.
↑ Cooke, J. A. L., Roth, V. D., and Miller, F. H. "The urticating hairs of theraphosid spiders". American Museum novitates (American Museum of Natural History) (2498). Retrieved 2008-10-11.
↑ Blackledge, T. A., and Wenzel, J. W. (2001). "Silk Mediated Defense by an Orb Web Spider against Predatory Mud-dauber Wasps". Behaviour 138 (2): 155–171.doi:10.1163/15685390151074357.
↑ Vollrath, F. (1986). "Eusociality and extraordinary sex ratios in the spider Anelosimus eximius (Araneae: Theridiidae)". Behavioral Ecology and Sociobiology 18 (4): 283–287.doi:10.1007/BF00300005.
↑ Agnarsson, I., and Kuntner, M. (2005). "Madagascar: an unexpected hotspot of social Anelosimus spider diversity (Araneae: Theridiidae)". Systematic Entomology 30 (4): 575–592.doi:10.1111/j.1365-3113.2005.00289.x.
↑ Avilés, L., Maddison, W. P. and Agnarsson, I. (2006). "A New Independently Derived Social Spider with Explosive Colony Proliferation and a Female Size Dimorphism". Biotropica 38 (6): 743–753. doi:10.1111/j.1744-7429.2006.00202.x.
↑ Matsumoto, T. (1998). "Cooperative prey capture in the communal web spider, Philoponella raffray (Araneae, Uloboridae)" (PDF).Journal of Arachnology 26: 392–396. Retrieved 2008-10-11.
↑ Bertani, R., Fukushima, C. S., and Martins, R. (2008). "Sociable widow spiders? Evidence of subsociality in LatrodectusWalckenaer, 1805 (Araneae, Theridiidae)". Journal of Ethology 26 (2): 299.doi:10.1007/s10164-007-0082-8.
↑ Meehan, C. J., Olson, E. J. and Curry, R. L. (21 August 2008). "Exploitation of the Pseudomyrmex–Acacia mutualism by a predominantly vegetarian jumping spider (Bagheera kiplingi)". 93rd ESA Annual Meeting. Retrieved 2008-10-10.
↑ Vollrath, F., and Knight, D. P. (2001). "Liquid crystalline spinning of spider silk". Nature 410(6828): 541–548. Bibcode:2001Natur.410..541V. doi:10.1038/35069000.PMID 11279484.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ Eberhard, W. G. (1990). "Function and Phylogeny of Spider Webs" (PDF). Annual Reviews in Ecology and Systematics 21: 341–372. doi:10.1146/annurev.es.21.110190.002013. Retrieved 2008-10-15.
↑ Agnarsson, I. (2004). "Morphological phylogeny of cobweb spiders and their relatives (Araneae, Araneoidea, Theridiidae)".Zoological Journal of the Linnean Society 141 (4): 447–626.doi:10.1111/j.1096-3642.2004.00120.x.
↑ RUPPERT, E.E., FOX, R.S. & BARNES, R.D. 2005. Zoologia dos Invertebrados.7ª ed., Ed. Roca, São Paulo.
↑ Coddington, J.A. 2005. Phylogeny and Classification of Spiders. In D. Ubick, P. Paquin, P. E. Cushing, and V. Roth (eds.) Spiders of North America: an identification manual, American Arachnological Society. 377 pages. Chapter 2, pp. 18-24.
↑ Norman I. Platnick. «The World Spider Catalog» (em inglês)
↑ Foelix, Rainer F. (1996). Biology of Spiders. 198 Madison Ave. NY, New York, 10016: Oxford University Press. p. 3. ISBN 0-19-509593-6.
↑ SANTOS, A. J.; BRESCOVIT, A. D.; JAPYASSÚ, H. F. Diversidade de aranhas: Sistemática, ecologia e inventários de fauna. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 1-23.
↑ SOUZA, A. L. T. Influência do habitat na abundância e diversidade de aranhas. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 24-43.
↑ Coddington, J. A. (2005). "Phylogeny and Classification of Spiders" (PDF). In Ubick, D., Paquin, P., Cushing, P. E., and Roth, V. Spiders of North America: an identification manual.American Arachnological Society. pp. 18–24. ISBN 0-9771439-0-2. Retrieved 2008-10-12.
↑ Coyle, F. A. (1986). "The Role of Silk in Prey Capture". In Shear, W. A. Spiders—webs, behavior, and evolution. Stanford University Press. pp. 272–273. ISBN 0-8047-1203-4.
↑ Penney, D. and Selden, P. A. Assembling the Tree of Life—Phylogeny of Spiders: a review of the strictly fossil spider families(PDF). In Deltshev, C., and Stoev, P. "European Arachnology 2005".Acta Zoologica Bulgarica. Supplement No. 1: 25–39. Retrieved 2008-10-13.
↑ Coddington, J. A. (2005). "Phylogeny and Classification of Spiders" (PDF). In Ubick, D., Paquin, P., Cushing, P. E., and Roth, V. Spiders of North America: an identification manual.American Arachnological Society. pp. 18–24. ISBN 0-9771439-0-2. Retrieved 2008-10-12.
↑ Meehan, C. J., Olson, E. J. and Curry, R. L. (21 August 2008). "Exploitation of the Pseudomyrmex–Acacia mutualism by a predominantly vegetarian jumping spider (Bagheera kiplingi)". 93rd ESA Annual Meeting. Retrieved 2008-10-10.
↑ Foelix, Rainer F. Biology of Spiders (2nd ed.). New York: Oxford University Press. pp. 4–5. ISBN 0-19-973482-8.
↑ Coddington, J. A. (2005). "Phylogeny and Classification of Spiders" (PDF). In Ubick, D., Paquin, P., Cushing, P. E., and Roth, V. Spiders of North America: an identification manual.American Arachnological Society. pp. 18–24. ISBN 0-9771439-0-2. Retrieved 2008-10-12.
↑ Coddington, J. A. (2005). "Phylogeny and Classification of Spiders" (PDF). In Ubick, D., Paquin, P., Cushing, P. E., and Roth, V. Spiders of North America: an identification manual.American Arachnological Society. pp. 18–24. ISBN 0-9771439-0-2. Retrieved 2008-10-12.
↑ "Natural history of Mygalomorphae". Agricultural Research Council of New Zealand. Retrieved 2008-10-13.
↑ SOUZA, A. L. T. Influência do habitat na abundância e diversidade de aranhas. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 24-43.
↑ SANTOS, A. J.; BRESCOVIT, A. D.; JAPYASSÚ, H. F. Diversidade de aranhas: Sistemática, ecologia e inventários de fauna. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 1-23.
↑ SANTOS, A. J.; BRESCOVIT, A. D.; JAPYASSÚ, H. F. Diversidade de aranhas: Sistemática, ecologia e inventários de fauna. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 1-23.
↑ SOUZA, A. L. T. Influência do habitat na abundância e diversidade de aranhas. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 24-43.
↑ SOUZA, A. L. T. Influência do habitat na abundância e diversidade de aranhas. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 24-43.
↑ SANTOS, A. J.; BRESCOVIT, A. D.; JAPYASSÚ, H. F. Diversidade de aranhas: Sistemática, ecologia e inventários de fauna. In: GONZAGA, M. O.; SANTOS, A. J.; JAPYASSÚ, H. F. (Orgs.) Ecologia e Comportamento de aranhas. Rio de Janeiro: Interciência, 2007. p. 1-23.
↑ «Ministério da Saúde»
↑ Salima Machkour M'Rabet, Yann Hénaut, Peter Winterton and Roberto Rojo (2011). "A case of zootherapy with the tarantula Brachypelma vagans Ausserer, 1875 in traditional medicine of the Chol Mayan ethnic group in Mexico". Journal of Ethnobiology and Ethnomedicine.
↑ Novak, K. (2001). "Spider venom helps hearts keep their rhythm".Nature Medicine 7 (155): 155. doi:10.1038/84588.PMID 11175840.
↑ Lewis, R. J., and Garcia, M. L. (2003). "Therapeutic potential of venom peptides" (PDF). Nature Reviews Drug Discovery 2 (10): 790–802. doi:10.1038/nrd1197. PMID 14526382. Retrieved 2008-10-11.
↑ Bogin, O. (Spring 2005). "Venom Peptides and their Mimetics as Potential Drugs" (PDF). Modulator (19). Retrieved 2008-10-11.
↑ Andrade E; Villanova F; Borra P; Leite, Katia; Troncone, Lanfranco; Cortez, Italo; Messina, Leonardo; Paranhos, Mario et al. (2008). "Penile erection induced in vivo by a purified toxin from the Brazilian spider Phoneutria nigriventer". British Journal of Urology International 102 (7): 835–7. doi:10.1111/j.1464-410X.2008.07762.x. PMID 18537953.
↑ Lisa Fritscher. «Spider Fears or Arachnophobia» (em inglês). Cópia arquivada em 2 de agosto de 2009
↑ 10 Most Common Phobias. «The 10 Most Common Phobias — Did You Know?» (em inglês). Cópia arquivada em 2 de agosto de 2009
↑ Friedenberg, J., and Silverman, G. (2005). Cognitive Science: An Introduction to the Study of Mind. [S.l.]: SAGE. pp. 244–245. ISBN 1412925681 !CS1 manut: Nomes múltiplos: lista de autores (link)
↑ Davey, G.C.L. (1994). «The "Disgusting" Spider: The Role of Disease and Illness in the Perpetuation of Fear of Spiders». Society and Animals. 2 (1). pp. 17–25. doi:10.1163/156853094X00045
↑ CAVALCANTE, Messias S. Comidas dos Nativos do Novo Mundo. Barueri, SP. Sá Editora. 2014, 403p.ISBN 9788582020364
↑ BASTOS, Abguar. A pantofagia ou as estranhas práticas alimentares da selva: Estudo na região amazônica. São Paulo, Editora Nacional; Brasília DF, INL. 1987, 153 p.
↑ REVISTA DE ATUALIDADE INDÍGENA. Aventuras de um índio que se tornou sertanista. p. 50-58. In: Revista de Atualidade Indígena. Brasília, Fundação Nacional do Índio. 1979, ano III, nº 19, 64 p
Ligações externas |
- aranhas.info - Página sobre as espécies de aranhas de Portugal
- Lista das espécies de aranhas de Portugal com fichas e fotos em Português
- Aranhas, acrobatas e perigosas - NAEQ - Núcleo de Apoio ao Ensino da Química
| Controle de autoridade |
|
|---|


