Espermatogênese
Espermatogénese | |
|---|---|
 | |
Um túbulo seminífero, com as células da linha germinativa. Coloração H&E. | |
 | |
Esquema de um espermatozóide humano maturo | |
Sistema | reprodutor masculino |
A espermatogénese (português europeu) ou espermatogênese (português brasileiro) é o processo fisiológico no qual se produzem os espermatozóides a partir de células germinativas. As células germinativas sofrem processo de divisão celular (mitose) uma e segunda vez formando as células-base cujo nome é espermatogónias (período de multiplicação) que, após crescimento sem divisão celular, dão origem aos espermatócitos primários (período de crescimento). Estes sofrem divisão meiótica (meiose I) e formam dois espermatócitos secundários; cada um destes espermatócitos secundários divide-se em dois espermatídeos (meiose II). Estes diferenciam-se, através de um processo chamado espermiogénese, em espermatozóides. Assim, cada espermatócito primário dá origem a duas células, os espermatócitos secundários, e os dois espermatócitos secundários dividem-se e diferenciam-se em quatro espermatozóides.
Os espermatozóides são os gâmetas masculinos em muitos organismos de reprodução sexuada. Deste modo, a espermatogénese é a vertente masculina da gametogénese, sendo essencial para a reprodução sexual. Nos mamíferos, ocorre nos túbulos seminíferos, nos testículos. A espermatogénese é altamente dependente de condições óptimas, nomeadamente a temperatura, para que ocorra correctamente. A metilação do DNA e a modificação das histonas têm um papel na regulação deste processo.[1] A espermatogénese tem início na puberdade, e, em circunstâncias normais, continua ininterruptamente até à morte, embora haja a tendência para a diminuição na quantidade de esperma produzido com o aumento da idade.
Índice
1 Localização
2 Duração
3 Etapas da espermatogénese
3.1 Espermatocitogénese
3.2 Espermatidogénese
3.3 Espermiogénese
4 O papel das células de Sertoli
5 Factores que influenciam a espermatogénese
6 Regulação hormonal
7 Ver também
8 Referências
Localização |
A espermatogénese ocorre em várias estuturas do sistema reprodutor masculino. As etapas iniciais têm lugar nos testículos, passando depois para o epidídimo, onde os gâmetas em desenvolvimento sofrem maturação e são armazenados até à ejaculação. Os túbulos seminíferos dos testículos são o ponto de partida do processo, onde as células basais adjacentes à parede interior do túbulo se dividem numa orientação centrípeta—começando nas paredes e progredindo até ao lúmen—produzindo espermatozóides imaturos. A maturação tem lugar no epidídimo.
A especificidade da localização destes processos é importante, uma vez que a espermatogénese requer uma temperatura mais baixa para produzir esperma viável (especificamente, uma temperatura de 1°-8 °C abaixo da temperatura normal do corpo, 37 °C).[2] Clinicamente, flutuações menores na temperatura, como aquelas induzidas pela utilização de coquilhas, não têm efeitos observáveis na viabilidade ou contagem de espermatozóides.[3]
Duração |
Nos seres humanos, o processo completo da espermatogénese demora 74 dias. Se se incluir o transporte no sistema de ductos, leva cerca de 3 meses. Os testículos produzem entre 200 e 300 milhões de espermatozóides diariamente.[4] Todavia, apenas metade se tornaram espermatozóides viáveis.[5]
Etapas da espermatogénese |
Todo o processo da espermatogénese pode ser dividido em várias etapas distintas, cada uma correspondendo a um tipo particular de célula nos seres humanos. Na tabela seguinte, os valores são dados para uma única célula, antes da síntese de DNA e da divisão (na G1, quando aplicável). O espermatócito primário é retido, entre a síntese de DNA e a divisão.
| Tipo celular | ploidia/cromossomas no ser humano | número de cópias de DNA/cromatídeos no ser humano | Etapa na qual a célula entra |
espermatogónia (tipos Ad, Ap e B) | diplóide (2N) / 46 | 2C / 46 | espermatocitogénese (mitose) |
espermatócito primário | diplóide (2N) / 46 | 4C / 2x46 | espermatidogénese (meiose I) |
| dois espermatócitos secundários | haplóide (N) / 23 | 2C / 2x23 | espermatidogénese (meiose II) |
| quatro espermatídeos | haplóide (N) / 23 | 1C / 23 | espermiogénese |
| quatro espermatozóides | haplóide (N) / 23 | 1C / 23 | espermiação |
Espermatocitogénese |

Diagrama representando o processo de espermatogénese e a evolução das células de espermatócitos primários, para espermatócitos secundários, para espermatídeos, e para espermatozóides.
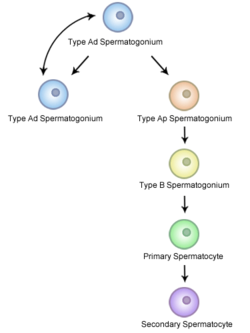
Diagrama esquemático da espermatocitogénese
A espermatocitogénese é a versão masculina da gametocitogénese, e resulta na formação de espermatócitos, com metade do material genético normal numa célula. Na espermatocitogénese, uma espermatogónia diplóide, presente no compartimento basal dos túbulos seminíferos, entra em divisão mitótica, dando origem a duas células intermédias diplóides chamadas espermatócitos primários. Cada espermatócito primário migra para o compartimento adluminal dos túbulos seminíferos, onde duplica o seu DNA e subsequentemente entra em meiose I, dando origem a dois espermatócitos secundários, haplóides, que mais tarde se dividem em espermatídeos haplóides. Esta divisão implica a ocorrência de recombinação genética, como são exemplos a inclusão aleatória dos cromossomas parentais, e o crossing-over, que aumentam a variabilidade genética do gâmeta.
Cada divisão celular (de espermatogónia para espermatídeo) é incompleta; as células permanecem ligadas uma à outra por pontes de citoplasma para permitir um desenvolvimento síncrono. Vale também a pena mencionar que nem todas as espermatogónias se dividem para produzir espermatócitos; se assim fosse, o número de espermatogónias reduzir-se-ia até à sua extinção. Pelo contrário, certos tipos de espermatogónia sofrem divisões mitóticas de modo a produzir cópias, assegurando um número de espermatogónias suficiente a permitir a espermatogénese continuamente.[6]
Espermatidogénese |
A espermatidogénese é a origem dos espermatídeos a partir dos espermatócitos secundários. Os espermatócitos secundários, produzidos nas etapas anteriores, entram rapidamente na meiose II, dividindo-se e originando espermatídeos haplóides.
A rapidez desta etapa é o motivo pelo qual os espermatócitos secundários são raramente vistos em estudos histológicos.
Espermiogénese |
Durante a espermiogénese, os espermatídeos começam por formar uma flagelo, pelo crescimento de microtúbulos num dos centríolos, que se torna no corpo basal. Estes microtúbulos formam um axonema. A porção anterior do flagelo (que toma a designação peça intermédia) torna-se mais grossa divido à acumulação de mitocôndrias em torno do axonema, de modo a assegurar a produção de energia. O DNA do espermatídeo torna-se altamente condensado. O DNA é compactado, numa primeira etapa, com proteínas nucleares específicas, que são posteriormente substituidas com protaminas durante a elongação do espermatídeo. A cromatina resultante, muito condensada, não tem capacidade de ser transcrita. O complexo de Golgi, que se encontra em redor do núcleo condensado, torna-se no acrossoma. Um dos centríolos da célula alonga, formando a cauda do espermatozóide.
Em seguida, tem lugar a maturação, que ocorre sob a influência da testosterona. Organelos e porções desnecessárias do citoplasma (os chamados corpos resíduais) são descartados, sendo fagocitados pelas células de Sertoli dos testículos. Os espermatozóides resultantes já maturaram mas não são dotados de motilidade, sendo que, nesta etapa, são ainda inviáveis. Os espermatozóides são libertados de entre as células de Sertoli para o lúmen do túbulo seminífero, num processo designado espermiação.
Os espermatozóides sésseis são transportados para o epidídimo com o fluido testicular secretado pelas células de Sertoli, com o auxílio de movimentos peristálticos. No epidídimo, os espermatozóides ganham motilidade e tornam-se capazes de fertilizar um oócito. Não obstante, o transporte dos espermatozóides maturos no restante sistema reprodutor masculino é feito por contracção muscular, e não pela motilidade própria do espermatozóide.
O papel das células de Sertoli |
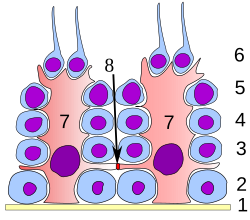
Representação esquemática da organização das células de Sertoli (a vermelho) e dos espermatócitos (a azul) no testículo. Os espermatídeos que ainda não sofreram espermiação estão ligados ao ápice luminal da célula.
Em todas as etapas de diferenciação, as espermatogónias estão em contacto próximo com células de Sertoli, que se julga servirem de apoio estrutural e metabólico às células da linha germinativa. Uma única célula de Sertoli estende-se desde a membrana basal até ao lúmen do túbulo seminífero, ainda que os processos citoplasmáticos sejam dificilmente discerníveis simplesmente ao nível microscópico.
As células de Sertoli desempenham uma série de funções durante a espermatogénese, e suportam os gâmetas em desenvolvimento das seguntes maneiras:
- Mantêm o ambiente necessário para o desenvolvimento e maturação, através da barreira hemato-testicular
- A barreira hemato-testicular é ainda importante na protecção dos espermatídeos do próprio sistema imunitário do indivíduo.
- Segregam substâncias iniciadoras da meiose
- Segregam fluido testicular
- Segregam proteína ligadora de andrógenos (ABP), que concentra a testosterona em proximidade aos gâmetas em desenvolvimento
- A testosterona é necessária em concentrações elevadas, para a manutenção do tracto reprodutivo, e a ABP permite um nível muito mais elevado de fertilidade
- Segrega hormonas que afectam a regulação hormonal da espermatogénese por parte da glândula pituitária, particularmente a inibina
- São células-alvo da folículo-estimulina (FSH)
- Fagocitose dos corpos residuais decorrentes da espermiogénese
- Segregam a hormona anti-Mülleriana, que inibem o desenvolvimento dos ductos de Müller[7]
As moléculas de adesão intercelular ICAM-1 e ICAM-1 solúvel têm efeitos antagonistas às zonulae occludentes que constituem a barreira hemato-testicular.[8] As moléculas de ICAM-2 regulam a adesão dos espermatídeos na porção apical da barreira (junto ao lúmen).[8]
Factores que influenciam a espermatogénese |
O processo da espermatogénese é altamente sensível a flutuações no ambiente, particularmente no que diz respeito à concentração hormonal e à temperatura. A testosterona tem que estar presente em elevadas concentrações locais para manter o processo, algo que é assegurado pela proteína ligadora de andrógenos presente nos túbulos seminíferos. A testosterona é produzida por células intersticiais, também chamadas de células de Leydig, que se encontram numa região adjacente aos túbulos seminíferos.
O epitélio seminífero é sensível a temperaturas elevadas nos seres humanos e nalgumas outras espécies, e exposição a temperaturas tão elevadas como a própria temperatura corporal pode surtir efeitos adversos. É por este motivo que os testículos se encontram for do corpo, no saco escrotal. A temperatura óptima para a espermatogénese é de 2°C (no homem)–8 °C (no rato) abaixo da temperatura corporal. A termorregulação faz-se através da regulação do fluxo sanguíneo[9] e da distensão/contracção dos músculos cremaster e dartos, que implica um afastamento/aproximação do testículo do resto do corpo.
Deficiências nutricionais (como a falta de vitamina B, E, e A), esteróides anabolizantes, metais (cádmio e chumbo), exposição a raios x, dioxinas, álcool, e doenças infecciosas podem afectar adversamente a taxa de espermatogénese. A linha germinativa é ainda susceptível a danos no DNA provocados pelo stress oxidativo que tem impactos significativos, provavelmente, na fertilização e na gravidez.[10] A exposição a pesticidas também afecta a espermatogénese.[11]
Regulação hormonal |
A regulação hormonal da espermatogénese varia de espécie para espécie. Nos seres humanos, o mecanismo não é completamente compreendido; todavia, sabe-se que a espermatogénese tem início na puberdade devido à interacção do hipotálamo, hipófise, e das células de Leydig. Se se tiver procedido à remoção da hipófise, a espermatogénese ainda se inicia, devido à folículo-estimulina e à testosterona.
A folículo-estimulina estimula tanto a produção da proteína ligadora de andrógenos pelas células de Sertoli, como a formação da barreira hemato-testicular. A proteína ligadora de andrógenos é essencial para manter a testosterona testicular em concentrações suficientemente elevadas para iniciar e manter a espermatogénese, que podem ser 20-50 vezes mais elevadas do que a concentração desta hormona no sangue. A folículo-estimulina pode iniciar a retenção da testosterona nos testículos, mas apenas a testosterona é necessária para manter a espermatogénese. Todavia, o aumento dos níveis da folículo-estimulina faz aumentar a produção de espermatozóides ao previnir a apoptose de espermatogónias de tipo A. A hormona inibina actua de modo a reduzir os níveis da hormona folículo-estimulina. Estudos em roedores sugerem que as gonadotopinas (tanto a LH como a FSH) auxiliam o processo espermatogenético ao suprimir os sinais pro-apoptóticos e, por conseguinte, ao promover a sobrevivência de espermatogónias.[12]
As células de Sertoli, por si só, mediam a espermatogénese através da secreção hormonal. São capazes de produzir as hormonas estradiol e inibina. As células de Leydig também produzem estradiol, para além da testosterona, o seu principal produto de secreção.
Ver também |
- Gametogênese
- Ovulogênese
- Fecundação
Referências
↑ Song, Ning; Liu, Jie; An, Shucai; Nishino, Tomoya; Hishikawa, Yoshitaka; Koji, Takehiko (2011). «Immunohistochemical Analysis of Histone H3 Modifications in Germ Cells during Mouse Spermatogenesis». Acta Histochemica et Cytochemica. 44 (4): 183–90. PMC 3168764 . PMID 21927517. doi:10.1267/ahc.11027
. PMID 21927517. doi:10.1267/ahc.11027
↑ "scrotum". Encyclopædia Britannica. Encyclopædia Britannica Online. Encyclopædia Britannica Inc., 2015. Web. 14 Jan. 2015 <http://www.britannica.com/EBchecked/topic/530078/scrotum>.
↑ Wang C, McDonald V, Leung A, Superlano L, Berman N, Hull L, Swerdloff RS (1997). «Effect of increased scrotal temperature on sperm production in normal men». Fertil. Steril. 68 (2): 334–9. PMID 9240266. doi:10.1016/s0015-0282(97)81525-7
↑ Padubidri, VG; Daftary, SN, eds. (2011). Shaw's Textbook of Gynaecology 15th ed. [S.l.: s.n.] p. 201. ISBN 978-81-312-2548-6
↑ Johnson L, Petty CS, Neaves WB (1983). «Further quantification of human spermatogenesis: germ cell loss during postprophase of meiosis and its relationship to daily sperm production». Biol. Reprod. 29 (1): 207–15. PMID 6615966. doi:10.1095/biolreprod29.1.207
↑ Fishelson, Lev; Gon, Ofer; Holdengreber, Vered; Delarea, Yakob (2007). «Comparative spermatogenesis, spermatocytogenesis, and spermato-zeugmata formation in males of viviparous species of clinid fishes (Teleostei: Clinidae, Blennioidei)». The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. 290 (3): 311–23. PMID 17525946. doi:10.1002/ar.20412
↑ Hadley, Mac E.; Levine, Jon E. (2007). Endocrinology 6th ed. Upper Saddle River, NJ: Prentice Hall. p. 369. ISBN 0-13-187606-6
↑ ab Xiao, X.; Mruk, D. D.; Cheng, C. Y. (2013). «Intercellular adhesion molecules (ICAMs) and spermatogenesis». Human Reproduction Update. 19 (2): 167–86. PMC 3576004. PMID 23287428. doi:10.1093/humupd/dms049
↑ Harrison, RG; Weiner, JS (1949). «Vascular patterns of the mammalian testis and their functional significance». The Journal of experimental biology. 26 (3): 304–16, 2 pl. PMID 15407652
↑ Lewis, SE; Aitken, RJ (2005). «DNA damage to spermatozoa has impacts on fertilization and pregnancy». Cell and tissue research. 322 (1): 33–41. PMID 15912407. doi:10.1007/s00441-005-1097-5
↑ Mehrpour, O; Karrari P (2014). «Occupational exposure to pesticides and consequences on male semen and fertility: A review.». Toxicol Lett. 230: 146–156. PMID 24487096. doi:10.1016/j.toxlet.2014.01.029|acessodata=requer|url=(ajuda)
↑ Pareek, Tej K.; Joshi, Ayesha R.; Sanyal, Amartya; Dighe, Rajan R. (2007). «Insights into male germ cell apoptosis due to depletion of gonadotropins caused by GnRH antagonists». Apoptosis. 12 (6): 1085–100. PMID 17268770. doi:10.1007/s10495-006-0039-3


